Как отмечалось выше, пол зародыша принципиально определяется при оплодотворении яйцеклетки. Яйцеклетка всегда содержит Х-хромосому, и пол зародыша зависит от того, какой сперматозоид ее оплодотворит—содержащий Х-хромосому (женский вариант) или Y-хромосому (мужской вариант). В женском организме одна из Х-хромосом подвергается спирализации, приводящей к образованию компактной глыбки полового хроматина. Наличие полового хроматина удается установить на 12—16-й день развития зародыша, если кариотип соответствует женскому. Интересно, что спирализации подвергается любая из Х-хромосом. Половина клеток женского организма имеет отцовскую Х-хромосому, а половина— материнскую, тогда как в мужском организме все Х-хромосомы имеют только материнское происхождение. Предполагается, что инактивация второй Х-хромосомы у девочек происходит не полностью и остаются локусы, ответственные за половое развитие. Предполагается также, что основная (неспирализованная) Х-хромосома ответственна за соматические процессы, но не играет существенной роли в половом диморфизме.
На ранних стадиях развития имеются две пары каналов для выведения гамет — Вольфовы протоки (производные мезонефроса) и Мюллеровы каналы (производные парамезонефроса). Благодаря классическим опытам Жоста (A. Jost, 1947, 1953) было установлено, что если гонада не развивается (или удалена), то со временем Вольфовы протоки подвергаются резорбции, а Мюллеровы каналы продолжают развиваться, превращаясь во внутренние женские половые органы (яйцеводы и матку). То же самое происходит в случае развития гонады по женскому типу. И только если происходит развитие семенника, Вольфовы протоки развиваются в семявыносящие канальцы, а Мюллеровы каналы подвергаются атрофии.
Впоследствии выяснилось, что развитие внутренних половых органов по мужскому типу обусловлено эмбриональной секрецией семенника и отсутствие таковой обязательно приводит к развитию женских половых признаков у плода, даже имеющего мужской набор половых хромосом. В то же время развитие гениталий у самки по женскому типу можно предотвратить путем введения андрогенов или подсадкой семенника.
Таким образом, гениталии всегда дифференцируются независимо от хромосомного набора клеток. Яичник зародыша не является источником гормонального воздействия на развитие половых органов.
Следующий важный момент, на котором следует остановиться, — формирование гипоталамо-гипофизарной системы регуляции половых желез. Известно, что гонадотропины у женщин выделяются циклически и эта цикличность связана с особенностями группы ядер гипоталамуса (преимущественно преоптической зоны). У лиц мужского пола наблюдается тонический тип выделения гонадотропинов. Благодаря опытам Грипа и Джона (O. Greep, L. Jones, 1950) было установлено, что гипофиз не обладает половыми различиями и, будучи пересаженным от самца к самке, начинает функционировать циклически.
Впоследствии выяснилось, что в эмбриогенезе существует критический период, во время которого семенник должен «сформировать» гипоталамус по мужскому типу с помощью андрогенов. Если беременной женщине в определенный период вводить большие дозы половых стероидов, то зародыш, даже при наличии женского кариотипа и яичников, будет обладать мужским типом секреции гонадотропинов.
Развитие самой гонады происходит в направлении формирования структур, изолирующих половые клетки от соматических, а также обеспечивающих половым клеткам возможность выхода во внешнюю среду. Основным различием в этом смысле является одиночный или групповой характер такой изоляции гоноцитов. Первый присущ яичнику (развитие фолликулов в корковой части гонады), второй — семеннику (семенные канальцы образуются из медуллярной закладки). Эти процессы детерминированы генетически и слабо контролируются путем гормонального воздействия. Однако после морфологического оформления гонады приобретают чувствительность к стимуляции гонадотропинами, хотя и не исключено влияние последних на число половых клеток, прошедших первичную стадию своего развития (до мейоза). Стромальные, клетки гонад очень чувствительны к стимуляции со стороны гипофиза. Так, мальчики рождаются с хорошо развитыми гландулоцитами, которые, однако, начиная с 7—14-го дня после рождения претерпевают обратное развитие. В последующем гландулоциты вновь начинают развиваться в период полового созревания.
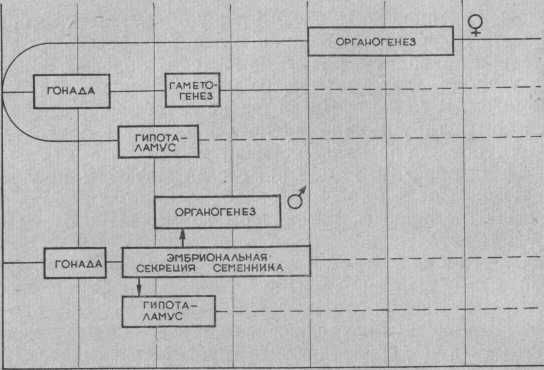
Рис. Принципиальная схема дифференцировки пола. Обозначения: I — девочки, II— мальчики.
Следует подчеркнуть следующие принципиальные моменты внутриутробной дифференцировки пола. Активность половых хромосом проявляется на очень коротком отрезке онтогенеза — в период дифференцирования пола гонады, причем их действие направлено лишь на ткань последней. Все остальные органы проходят стадию развития независимо от половых хромосом. Развитие гипоталомогипофизарных структур, а также внутренних и наружных половых органов генетически дифференцировано по женскому типу, и лишь эмбриональная инкреция семенника способна изменить их развитие на развитие по мужскому типу.