В процессе полового развития период полового созревания (т. е. период, когда появляются вторичные половые признаки) занимает особое место. Происходящие здесь изменения охватывают без исключения все органы и системы организма, однако главное, к чему можно свести этот процесс, — созревание репродуктивной системы.
Зависимость деятельности половых желез от гипофиза была установлена в начале XX в. В последующем из гипофиза были выделены три гормона, стимулирующие репродуктивную систему: фоллитропин (фолликулостимулирующий), лютропин (лютеинизирующий) и пролактин. Пролактин способствует лактации и молокоотделению. В то же время пролактин не имеет никакого отношения к росту молочных желез. Напротив, он резко снижает число митозов в их железистом эпителии. Этим отчасти объясняется тот факт, что у женщин, длительно кормивших грудью, рак молочных желез встречается реже, чем у не кормивших. Также очевидно, что количество молока, выделяющегося при кормлении, определяется не величиной железы, а интенсивностью ее стимуляции со стороны гипофиза. У мужчин биологическая роль пролактина остается невыясненной.
Фоллитропин и лютропин играют существенную роль в деятельности гонад. Удаление гипофиза прекращает выработку гормонов и половых клеток в гонадах, приводит к атрофии последних. В тех случаях, когда атрофия не зашла слишком далеко, заместительная терапия восстанавливает утраченную функцию половых желез.
У женщин в течение менструального цикла в механизм нейрогуморальной регуляции последовательно включаются все три гонадотропных гормона. Фоллитропин стимулирует рост и созревание фолликулов в яичнике, а также образование эстрогенов. Вместе с тем, для того чтобы половые гормоны вырабатывались зрелым фолликулом в оптимальном количестве, необходимо одновременное воздействие и лютропина. Последний способствует образованию и развитию желтого тела. Секреция прогестерон на желтым телом происходит под влиянием одновременного воздействия лютропина и пролактина.
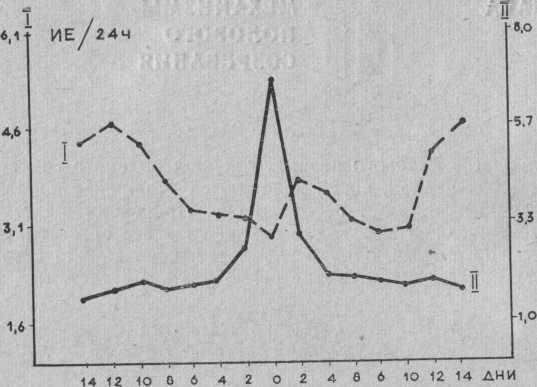
Рис. Выделение фоллитропина (I) и лютропина (II) с мочой у женщин на протяжении овариального цикла (за 0 принят день овуляции).
Лютропин обнаруживается в моче женщин во всех стадиях менструального цикла; нарастание уровня его выделения начинается до овуляции. На протяжении двухфазного менструального цикла в фолликулиновую фазу экскреция лютропина находится в пределах 1— 2 ИЕ/24 ч, ко времени овуляции она повышается на 7—8 ИЕ/24 ч. Кривая экскреции фоллитропина имеет два повышения, в принципе соответствующие пикам экскреци эстрогенов. Вместе с тем максимум выделения эстрогеноц и гонадотропинов перед овуляцией не всегда совпадает; иногда пик выделения гонадотропинов, наступает на 1—4 дня позже пика эстрогенов, но не наоборот.
Из гормонов задней доли гипофиза в регуляции менструального цикла участвует окситоцин. Кроме непосредственного воздействия на гладкую мускулатуру матки оксптоцин способствует овуляции и секреции пролактина. Выделяясь во время кормления грудью, он усиливает выброс молока грудной железой. В связи с этим женщины во время кормления грудью нередко ощущают сокращение матки.
В мужском организме лютропин стимулирует секрецию андрогенов интерстициальными клетками яичек. Введенный в организм, он практически не влияет на сперматогенез. Так же как и у женщин, фоллитропин, преимущественно гаметокинетический фактор, стимулирует выработку половых клеток, однако он более эффективен в присутствии лютропина. Таким образом, для стимуляции гонад требуется совместное воздействие фоллитропина и лютропина.
Предположение, что функция гипофиза регулируется особыми центрами в гипоталамусе (W. Hohlweg, 1934), было подтверждено впоследствии Харрисом (G. Harris, 1955), который доказал несомненную зависимость гонадотропной функции гипофиза от деятельности срединного возвышения (eminentia mediana) гипоталамуса. В эксперименте введение экстрактов, полученных из этой области межуточного мозга, приводит к усиленному выбросу гонадотропинов из гипофиза и повышению их уровня в периферической крови. Вещества, выделяемые нейронами гипоталамуса и стимулирующие функцию гипофиза, получили название либеринов (релизинг-факторов), т. е. факторов, «освобождающих» гонадотропины.
В настоящее время известны либерины для всех гормонов передней доли гипофиза, включая и указанные выше гонадотропные гормоны. Эти соединения относятся к классу коротких пептидов с относительно низким молекулярным весом (для фоллиберина он составляет около 300). Каждый Либерии обладает относительно высокой специфичностью, за исключением, пожалуй, люлиберина, который стимулирует выброс и лютропина и фоллитропина (правда, влияние люлиберина на выделение фоллитропина на 20—30% меньше, чем фоллиберина).
Ядра гипоталамуса обладают не только тропной, но и половой специфичностью. Благодаря работам Барклоу и Горского (С. Barraclough, R. Gorski, 1961) было установлено, что аркуатные ядра ответственны за тоническое выделение гонадотропинов, характерное для самцов, тогда как у самок гонадотропная функция гипофиза регулируется преимущественно супраоптическими и паравент рикулярными ядрами; последние стимулируют циклическое выделение гонадотропинов.
Кастрация у взрослых быстро приводит к повышению содержания гонадотропинов в гипофизе и периферической крови. В отдаленные сроки наблюдается базофильная гиперплазия передней доли гипофиза. С другой стороны, при введении кастратам достаточных доз половых стероидов уровень гонадотропинов в плазме и гипофизе падает. Экзогенные стероиды способны подавлять продукцию гонадотропинов и у нормально развитых людей. Таким образом, существует отрицательная обратная связь между гипофизом и гонадами. Увеличение продукции гонадотропинов стимулирует повышенную выработку гонадами половых стероидов, усиленная же выработка последних приводит к снижению активности гипофиза. Однако этим дело не ограничивается.
В своих опытах Барклоу и Горский (1961) показали, что наиболее эффективно половые гормоны подавляют секрецию гонадотропинов в случае, если попадают непосредственно в гипоталамическую область. Тот факт, что механизм обратной связи замыкается на гипоталамусе, подтверждается следующим опытом: если стимулировать электрическим током ядра гипоталамуса, наблюдается выброс лютропина, введение же тестостерона блокирует этот процесс.
Регуляторные связи репродуктивной системы усложняются двойным назначением гонады. Так, в мужском организме секреция лютропина в общем находится в обратной зависимости от концентрации в крови тестостерона. Однако продукция фоллитропина снижается лишь при введении нефизиологически больших доз тестостерона либо при длительном его применении. В большей степени секреция фоллитропина коррелирует с интенсивностью последних стадий сперматогенеза. Многие авторы допускают, что сигнал об оптимальном уровне фоллитропина для сперматогенеза поступает из цитоплазмы сперматид в виде «Х-гормона» (ингибина).
Оценка характера взаимосвязи гипофиза с гонадами осложняется тем, что выработка гормонов и выделение их в кровь в значительной степени проявляют противоположные тенденции. Именно с этим связаны различия в эффекте разных доз стероидов. Так, эстрогены, тормозя продукцию гонадотропинов, могут усиливать их выделение из гипофиза. Если искусственно подавлять гонадотропную функцию гипофиза с помощью половых стероидов, то после отмены последних уровень гонадотропинов быстро восстанавливается и даже наблюдается его увеличение по сравнению с исходным. На этом принципе основан широко используемый в клинике метод лечения некоторых видов бесплодия.
Таким образом, в регуляции репродуктивной деятельности выявляются три типа обратной связи. Первый (длинная обратная связь) включает периферические железы и гипоталамус и осуществляет поддержание адекватного уровня гонадотропной активности с помощью половых гормонов. Второй (короткая обратная связь) включает гипофиз и гипоталамус. Наличие такой связи подтверждается тем, что экзогенное введение гонадотропинов приводит к снижению продукции в гипоталамусе фоллиберина и люлиберина и уменьшению содержания гонадотропинов в гипофизе. В последние годы предполагают существование третьего типа связи (ультракороткая обратная связь) непосредственно между гипоталамическими ядрами.
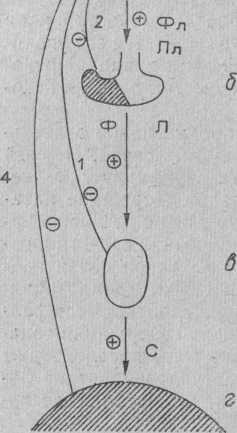
Рис. Основные звенья репродуктивной системы и пути ее регуляции. Обозначения: 1 — длинная обратная связь, 2 — короткая, 3 — ультрако-роткая, 4 — сверхдлинная; Фл — фоллиберин, Лл — люлиберин, Ф — фоллитропин, Л — лютропин, С — половые стероиды; Ч — стимулирующее влияние, тормозящее влияние; а — гипоталамус, б — гипофиз, в — гонады, г — гениталии и органы-мишени.
Таким образом, прямое регулирующее влияние на ре-продуктивную систему начинается с гипоталамуса и осуществляется по всем звеньям системы. Обратные импульсы, также поступают в гипоталамус, однако, возможно, минуя непосредственно вышестоящие звенья. Гипоталамус как бы обрабатывает и суммирует сигналы о состоянии всех звеньев системы, «принимает решение» и отдает директиву о целесообразном уровне функционирования нижележащих структур.